斯维特拉娜N. Radyuk
研究副教授
生物科学
| 办公室的位置 |
DLSB 340 |
| Phone |
214-768-2892 |
教育
Ph.D. 应用微生物研究所电话号码:214-768-2065
电子邮件: snradyuk@julihui168.com
教育
|
1979: |
化学,理科硕士.V.罗蒙诺索夫莫斯科国立大学,俄罗斯 |
|
1990: |
俄罗斯应用微生物研究所生物学博士 |
教学
免疫生物学(BIOL 4319)
分子遗传学实验室(BIOL 3222)
分子遗传学研究的概念(BIOL 6222)
生物科学概念(BIOL 6314)
生物学研究(BIOL 6370)
研究生研究(论文研究)(BIOL 6270)
论文(研究)(BIOL 8398)
研究兴趣
免疫与衰老
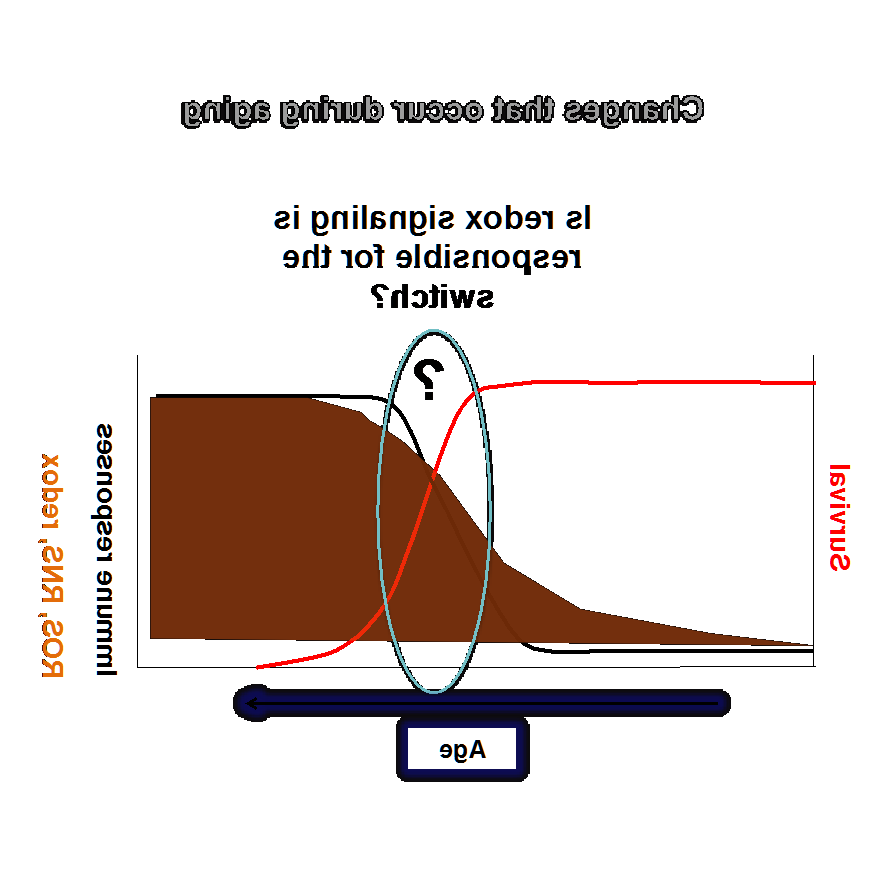
我们实验室目前的研究重点是了解氧化还原在先天免疫和衰老中的作用. 衰老的特征是免疫系统失灵. 在衰老过程中,免疫反应的放松会导致癌症和其他与年龄有关的疾病. 我们小组的研究旨在阐明这种放松管制的机制,以便制定适当的干预措施.
与衰老相关的最显著变化涉及免疫力:
•免疫反应能力下降
•对传染病和癌症的易感性增加
•降低对疫苗接种的反应
•T细胞和B细胞功能受损和克隆扩增
低水平的toll样受体
•老年生物体先天免疫增强,导致过度免疫反应和炎症
•延长寿命与先天免疫的最佳功能相关, 减少促炎反应和减少病原体负担
•增强的病原体抗性是具有长期生存背景的菌株的特征
大型生物对微生物挑战的反应和抵抗是有代价的:
•增强的免疫力提供了抵御入侵者的保护, 它也可能引起宿主细胞和组织的损伤, 最终会导致功能性缺陷吗.
•免疫效应物的过量产生也会对细胞机制产生大量的能量需求, 哪些可能导致对维持细胞内稳态产生负面影响的权衡.


免疫反应的氧化还原调控:
活性氧和活性氮(ROS和RNS), 在代谢反应中产生, 炎症, 和吞噬作用, 不仅会造成组织损伤, 还在信号通路中充当信使, 包括免疫反应的调节.
细胞氧化还原的主要调节因子之一是被称为硫醇依赖性过氧化物酶的酶, 或抗氧化蛋白. 它们是:
•广泛分布于三国
•在所有生物中高度保守,从细菌开始延伸到人类
•参与许多氧化还原相关的细胞功能:分化, 扩散, 细胞凋亡, 信令流程, 免疫反应.
目的是确定如何维持“正确”的ROS/RNS水平,以尽量减少组织损伤,同时不损害免疫反应. 我们的研究小组正在测试一种假设,即过氧化物还毒素可能控制氧化还原状态下的“开关”,从而调节免疫反应和衰老. 为了解决这些问题,我们正在使用广泛的分子生物学和遗传学技术.
实际研究两者中调节免疫和衰老的氧化还原信号的差异 young and old organisms; develop interventions, 考虑到对年轻人和老年人的不同影响.
目前支持
R01 AG032342 2011年4月1日- 2016年3月31日(首席研究员)
过氧化物还原素、免疫信号和衰老
由NIH/NIA资助
本研究的目的是确定过氧化物还毒素在调节免疫和衰老之间的关系中的作用,并确定确定这种关系的信号通路
R01 AG045830 2013年8月1日- 2018年5月31日(CoPI, PI -博士. Jadwiga Giebultowicz,俄勒冈州立大学)
“生物钟与衰老”
由NIH/NIA资助
这项研究的目的是确定氧化还原之间的功能联系, 强烈的外围生物钟和衰老的速度
选定的出版物
Klichko V.Sohal B.H.Radyuk年代.N., Orr W.C.索哈尔·R.S. (2014)细胞色素c氧化酶储备能力的降低降低了黑胃果蝇的健壮性,缩短了寿命. 物化学J. 2014年4月1日;49 (1):127-35.
Orr, W.C.Radyuk年代.N.索哈尔·R.S. (2013)氧化还原状态在果蝇衰老中的作用. 抗氧化氧化还原信号. 2013年9月10日;19(8):788-803.
Radyuk年代.N.克里奇科V.I.迈克尔·K., Orr W.C. (2013)过氧化氧还蛋白4对果蝇生理的影响是一个复杂的抗氧化和信号功能的相互作用. 美国实验生物学学会联合会J. 2013年4月,27 (4):1426 - 38.
海狸L.M.克里奇科V.I.,周娥.S., Kotwica-Rolinska J.威廉森·M., Orr W.C.Radyuk年代.N.[J].M. (2012)果蝇谷胱甘肽水平和生物合成的昼夜节律调节. 《威尼斯人娱乐城》. 2012;7(11):e50454. doi: 10.1371 /杂志.pone.0050454. 2012年11月30日.
Radyuk,年代. N.甘比尼,J.博拉斯,C.,塞尔娜,E.克里奇科,V. I., Viña, J., Orr, W. C. (2012)过表达谷氨酸半胱氨酸连接酶的长寿果蝇转录谱的年龄依赖性变化机制. 老化的重击. 2012年6月,133(6):401 - 13所示.
Radyuk年代N, Rebrin I克里奇科VISohal BH迈克尔·K贝内斯·J索哈尔·RS, Orr WC. (2010)线粒体过氧化物还毒素对于维持氧化还原状态和果蝇成虫的存活至关重要. 自由基生物医学. 49: 1892-1902.
Radyuk,年代. N.,雷布林,我.卢恰克,J. M.麦克拉克,K.克里奇科,V. I.Sohal, R. S., Orr, W. C. (2008). 果蝇谷氨酸-半胱氨酸连接酶的催化亚基是一种核细胞质穿梭蛋白. J. Biol. Chem. 2009年1月23日;284(4):2266-74.
Radyuk年代N, Rebrin I克里奇科VISohal BH迈克尔·K贝内斯·J索哈尔·RS, Orr WC. (2010)线粒体过氧化物还毒素对于维持氧化还原状态和果蝇成虫的存活至关重要. 自由基生物医学. 49: 1892-1902.
Radyuk,年代. N.麦克拉克,K.克里奇科,V. I., Benes, J., Orr, W. C. (2010). 过氧化氧还蛋白5调节果蝇的免疫反应. Biochim. et Biophys. 学报1800:1153-1163.
Radyuk,年代. N.麦克拉克,K.克里奇科,V. I., Benes, J.,雷布林,我.Sohal, R. S. Orr, W. C. (2009). 过氧化氧还蛋白5可以防止氧化应激和细胞凋亡,并促进果蝇的寿命. 物化学J. 419(2): 437-445.
L:., S. 拉杜克,T. A. 哈顿(2009). 用于生物防御和生物威胁监测的功能性磁性纳米颗粒, 分析化学, 81, 5637-5645.
Legan,年代. K.,雷布林,我.莫凯特,R. J.Radyuk,年代. N.克里奇科,V. I.Sohal, R. S., Orr, W. C. (2008). 葡萄糖-6-磷酸脱氢酶的过度表达延长了黑腹果蝇的寿命. 生物化学. 283:32492-32499.
的旧事,K., Orr, W. C.Radyuk,年代. N. (2008). 果蝇过氧化物还氧蛋白5是双双子操纵子中的第二个基因. 物化学. Biophys. Res. Commun. 368:273-278.
Luchak J. M.L . Prabhudesai.Sohal, R. S.Radyuk,年代. N.Orr, W. C. (2007)谷氨酸-半胱氨酸连接酶的过表达和过低表达对果蝇寿命的调节. Ann. N.Y. Acad. Sci. 1119:260-273.
Radyuk,年代. N.麦克拉克,K.,雷布林,我.Sohal, R. S., Orr, W. C. (2006). 果蝇DNA糖基酶dOgg1和RpS3在线粒体中异位表达的影响. 自由基生物医学. 41: 757-764.
Orr, W. C.Radyuk,年代. N.L . Prabhudesai.D .托罗泽., Benes, J. J.卢恰克,J. M.莫凯特,R. J.,雷布林,我.哈伯德,J. G. Sohal, R. S. (2005)谷氨酸-半胱氨酸连接酶的过度表达延长了黑腹果蝇的寿命. J. Biol. Chem. 280: 37331-37338.
Radyuk,年代. N.克里奇科,V. I. Orr, W. C. (2004)剖析Cu,锌-超氧化物歧化酶在果蝇中的表达-编码域内内含子/外显子序列的关键调控作用. 基因,17:328-48.
Klichko V. I.Radyuk,年代. N. Orr, W. C. (2004)果蝇发育和衰老过程中过氧化氢酶基因表达谱分析. Arch. 昆虫生物化学. 杂志. 56: 34-50.
Radyuk,年代. N.默里科,P. A.波波娃,T. G.格林,E. 阿里贝克,K. (2003)体外生成的呼吸道粘膜:研究吸入性炭疽的新工具. 物化学. Biophys. Res. Commun. 305: 624-632.
Radyuk年代. N.索哈尔·R. S. 和Orr W. C. (2003)硫氧还蛋白过氧化物酶可以促进细胞保护或细胞死亡,以应对不同的应激源——果蝇细胞中硫氧还蛋白过氧化物酶过表达和过表达的研究. 物化学. J. 371: 743-752.
Mockett R. J.Radyuk年代. N.贝内斯·J. J., Orr W. C. Sohal, R. S. (2003)家族性肌萎缩性侧索硬化症突变体Sod等位基因在转基因果蝇中的表型效应. Proc. Natl. Acad. Sci. U.S.A. 100: 301-306.
Radyuk,年代. N.克里奇科,V. I.斯皮诺拉,B.Sohal, R. S. Orr, W. C. (2001)果蝇过氧化物还氧蛋白基因家族. 自由·拉迪奇. Biol. Med. 31: 1090-1100.
Radyuk,年代. N.克里奇科,V. I. Orr, W. C. (2000)黑腹果蝇过氧化氢酶的表达对蜕皮激素有反应,并表现出转录和转录后调控. Arch. 昆虫生物化学. 杂志. 45: 79-93.
Klichko V. I.Radyuk,年代. N. Orr, W.C. (1999) CuZn-SOD启动子驱动在果蝇中枢神经系统中的表达. 一般人. 老龄化,20:537-543.
Mockett R. J., Orr, W. C.拉赫曼达,J. J., Benes, J.J.Radyuk,年代.N.克里奇科,V. I. Sohal, R. S. (1999)转基因黑腹果蝇含锰超氧化物歧化酶的过表达. Arch. 物化学. Biophys. 371: 260-269.
》,米.Sohal, R. S.Radyuk,年代. N.克里奇科,V. I. Orr, W. C. (1997)黑腹果蝇谷胱甘肽还原酶基因的分子结构. Arch.物化学.Bioph. 339: 323-334.
